Agencies of Pollination
In pollination process, the non-motile pollen grains are required to be transferred from anther to the sigma of flower.
For this puipose plants have to depend upon certain external carriers. Such cariers are described as agencies of pollinaion or pollinating agents.
Various agencies of pollinaion agents broadly classified in two major categories.
Abiotic agents: These are the nonliving or physical factors like wind or water which help in transfer of pollen grains from anther to the stigma of flower.
Biotic agents: These are the living organisms like insects, birds, bats etc. which help in transfer of pollen grains.
On the basis of agents involved, pollination is of following types:
Anemophily (Anemone = wind, phily = loving or attracting):
The transfer of pollen grains through wind is described as anemophily.
The wind pollinated plants are called anemophilous plants.
Anemophily is considered as the most primitive type of pollination and mostly the plants produce unisexual flowers.
Anemophilous plants make certain adaptations in their flowers to favour this process. These are called floral adaptations for anemophily.
Anemophilous flowers are small, inconspicuous without bright colours, nectar and fragrance. Petals are green or highly reduced.
When flowers are unisexual, the male flowers are more numerous than female and are situated at a higher level.
Stamens are with long filaments, versatile and exposed anthers.
Pollen grains are produced in extremely large numbers to compensate their wastage.
These are small, smooth walled, pale yellow in colour and light weighted.
Long style bears large feathery or hairy stigma to trap pollen grains. E.g. Grasses, Maize, Jowar, Sugarcane, etc.
Hydrophily:
The transfer of pollen grains through the agency of water is Hydrophily.
The water pollinated plants are called hydrophilous plants. Some of the aquatic plants make excellent use of water for their pollination.
Floral adaptations made by the hydrophilous plants are that their flowers are mostly unisexual, small and inconspicuous.
They are without fragrance, bright colour and nectar. Pollen grains and other floral parts are unwettable i.e. coated with mucilage.
Specific gravity of pollen grain is controlled by the formation of one or more starch grains in the pollen grain.
Stigma is long and sticky. Hydrophily is of two types; Hypohydrophily and Epihydrophily.
Hypohydrophily:
Pollination with the help of water takes place below the water surface in the hydrophytes bearing submerged female flowers.
Such plants produce needle like pollen grains which lack the exine.
They bear same specific gravity as that of water; therefore, they float below the water surface.
Such pollen grains when come in contact with stigma, they coil around it and germinate.
E.g. Zostera and Ceratophyllum
Epihydrophily
When pollination occurs on the surface of water, it is called epihydrdphily.
It is a more common type of hydrophily. Vallisneria is a dioecious plant i.e. male and female flowers are submerged and produced on separate plants.
At the time of maturity the male flowers are detached from the male inflorescence and begin to float on water surface.
The female flowers have a coiled, long pedicel, which undergoes uncoiling at the time of maturity.
They now reach the water surface. The male flowers surround the female flower and undergo anthesis (formation of pollen grains, at maturity) due to which pollen grains are deposited on the stigma of female flower and cross pollination is achieved.
Entomophily: (Entomon = insect)
Pollination through the agency of insects is called entomophily and these plants are called entomophilous.
Common insects which act as carriers in entomophily are moths, butterflies, beetles, flies, bees etc.
Bees are most common agents and nearly 75-80% of total insect pollination is achieved with the help of bees.
Entomophilous flowers are large and attractive, when small; flowers are produced in compact group or inlorescence e.g. Sunflower.
Flowers have attractive bright colors, pleasant fragrance and nectar.
In some plants, additional modifications are made for attraction of insects for e.g.. Corona in passion flower, petaloid bracts in Bougainvillea etc.
The pollen grains have spiny or rough exine.
Stigma is sticky. E.g. Jasmine, Rose, Cestrmn, Salvia, Bougainvillea.
Few plants develop special mechanism to favour this process e.g. Salvia has developed lever
In Salvia, flower is bisexual and protandrous.The two stamens of flower have long bifurcated (divaricate) connective.
Upper branch of connective bears fertile anther lobe while the flower has sterile anther lobe.
An insect when enters in the flower, it pushes the lower sterile lobe as a result the upper fertile anther lobe bends down.
It comes in contact with back side of insect body and pollen grains are dusted there.
The same insect when visits another flower with matured gynoecium, the pollen grains are picked up by the receptive stigma. Such mechanism is called lever - mechanism
Ornithophily - (Ornis = bird)
When the pollination takes place through the birds, then it is called ornithophily.
Such bird pollinated plants are called omithophilous plants. Ornithophilous plants show large flowers with thick and fleshy floral parts.
Corolla is tubular or funnel shaped. Flowers with bright colored corolla like orange, red, yellow, blue which help in attracting the birds.
Flowers lack fragrance because birds have poor sense of smell.
Flowers produce large amount of sugary nectar, which is used as a drink by the birds.
Pollen grains are sticky. E.g. Callistemon (Bottle brush), Bignonia, Butea and Bombax (Silk Cotton).
The common pollinating birds are; sunbirds, humming birds, crow, bulbul.
Chiropterophily- (Chiropteron = bat)
Pollination which takes place with the help of bats, is called chiropterophily and such plants are called chiropterophilous plants.
In these plants flowers are large, stout enough so that bats can hold on to the flowers.
Chiropterophilous plants ae nocturnal and open their flowers during night time.
Flowers emit rotten fruits like fermenting fruity odor.
Flowers produce copious nectar.
Flowers have large number of stamens to produce a considerably large quantity of pollen grains.
E.g. Anthocephallus (Kadamb), Kigellia pinata, Adansonia (Baobab tree), Bauhinia.
Out breeding Devices for Cross Pollination
Most of the Angiosperms produce bisexual flowers.
Cross pollination is preferred by majority of flowering plants and they develop out breeding devices.
In many plants self pollination is avoided as stamens and carpels do not mature simultaneously. This is called dichogamy.
In some plants, pollen grains are released much before stigma becomes receptive, this is protandry e.g. Sunlower.
In some plants, stigma becomes receptive first, this is protogyny e.g. Michelia.
The third device to avoid self pollination is genetic mechanism and it is called self- incompatibility or self sterility, e.g. Orchids. Pollen germination on the stigma of the same lower is inhibited.
The fourth device is to produce unisexual flowers,i.e. unisexuality.
In some plants like Castor, male and female flowers are produced on the same plant and such plants are called monoecious. In dioecious plants like Papaya, male and female flowers are produced on different plants. This condition prevents autogamy as well as geitonogamy while monoecious condition prevents only autogamy.
Pollen -Pistil Interaction:
All the events from deposition of pollen grain on the stigma to the entry of pollen tube in the ovule are referred to as pollen- pistil interaction.
It is very dynamic process involving pollen recognition followed by promotion or inhibition of pollen germination.
Cross pollination by any agent is always by chance. It does not guarantee the transfer of the right type of pollen i.e. compatible pollen of the same species as the stigma.
Quite often, the wrong type of pollen i.e. either from other species or from the same plant but it is self-incompatible, lands on the stigma.
The pistil has the ability to recognize the pollen. If it is right type, the pistil accepts it and promotes post-pollination events that lead to fertilization.
If it is wrong type, the pistil rejects it by preventing germination of pollen on the stigma or growth of pollen tube in the style.
Thus, only right type, compatible pollen, germi-nates on stigma and a pollen tube comes out through one of the germ pores.
The contents of the pollen grain move into the pollen tube and it grows through the tissues of stigma and then style.
In most of the plants, pollen grains are shed at the two celled stage (a vegetative cell and a generative cell), the generative cell divides and forms two male gametes during the growth of the pollen tube.
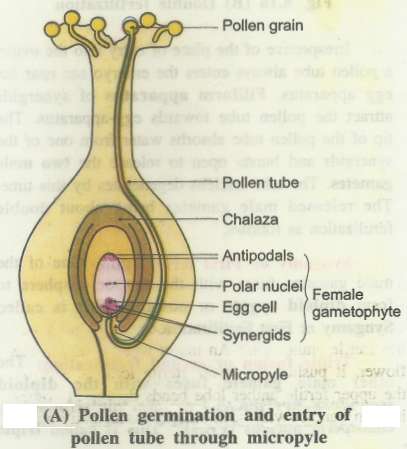
After reaching the ovary, pollen tube enters the ovule through the micropyle and then enters one of the synergids through the filiform apparatus and bursts to release male gametes.
All these events are part of pollen-pistil interaction. The ability of pistil to recognize the pollen is due to certain chemical components present in the pollen grain which interact with those of the pistil.
Recently, botanists have been able to identify some of the pollen and pistil components and the interactions leading to the recognition, followed by acceptance or rejection.
The knowledge gained in this area would cetainly help the plant breeders in manipulating pollen-pistil interaction, even in incompatible pollinations, to get desired hybrids.
DOUBLE FERTILIZATION: PROCESS AND SIGNIFICANCE:
" The process of fusion of male gamete with the female gamete to form the diploid zygote (2n) is known as fetilization"
The fusion of one male gamete with egg and that of another male gamate with secondry nucleus is called double fertilization.
All angiosperms are characterized by a unique process called "Double-Fertilization".
It was discovered by S.G. Nawaschin (1898) in Lillium and Fritillaria Species.
It occurs as follows:
After pollination, pollen grain germinates on the stigma to form a pollen tube.
Pollen tube grows down towards the ovule in ovary.
It carries two non motile male gametes, thin cytoplasm and a degenerating tube nucleus with it.
Growth of the pollen tube occurs at its tip where most of the cytoplasm is concentrated.
On reaching the ovary, a pollen tube mostly enter in an ovule through micropyle (porogainy).
Sometimes it may enter through integuments (mesogamy) or through chalaza (chalazogamy).
Irrespective of the place of entry into the ovule, a pollen tube always enters the embryo sac near the egg apparatus.
Filiform apparatus of synergids attract the pollen tube towards egg-apparatus.
The tip of the pollen tube absorbs water from one of the synergids and bursts open to release the two male gametes.
The tube nucleus degenerates by this time. The released male gametes bring about double fertilization as follows
1. Syngamy or First fertilization
2. Triple fusion or second fertilization
1. Syngamy or First fertilization
One of the male gametes fuses with the egg or oosphere to form diploid zygote or oospore. This is called Syngamy or First Fertilization.
2. Triple fusion or second fertilization
The other male gamete fuses with the diploid secondary nucleus to form the triploid primary endosperm nucleus (PEN).
This is called triple fusion. Since each of the polar nuclei is sister nucleus of the egg, it is called second fertilization.
Significance of Double Fertilization Process
The fertilization process in which non motile male gametes are transported up to the female gamete through a pollen tube is called Siphonogamy.
Double Fertilization is significant because it involve the use of both the male gametes produced by a pollen grain.
This avoids the chances of polyembryony and increases the chances of survival of future plant.
It consists of two fusions and results in formation of two products.
First fertilization restores diploidy in the life cycle. Diploid zygote develops into embryo which subsequently develops into new plant.
Second fertilization product, triploid PEN (Primary Endosperm Nucleus) develops to form a nutritive tissue called endosperm for developing embryo.
As the development of embryo and endosperm takes place close to each other, proper nutrition of embryo is ensured.
Development of embryo and endosperm takes place simultaneously; therefore, unnecessary wastage of endosperm is avoided.
The triploid endosperm gives better nutrition to the developing embryo; therefore, seeds are more viable in Angiosperms.
In Gymnosperms, multicellular, haploid, female gametophyte nourishes the developing embryo and is called endosperm.
Seeds are produced but there is no double fertilization. Thus it is the characteristic feature of only Angiosperms.
